Комплекс потенциалов мозга, связанных с движениями
|
|

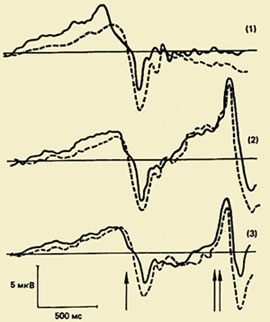 Одним из важных направлений в исследовании психофизиологии двигательного акта является изучение комплекса колебаний потенциалов мозга, связанных с движениями (ПМСД). Значение этого феномена для понимания физиологических механизмов организации движения очень велико, потому что изучение ПМСД позволяет выявить скрытую последовательность процессов, происходящих в коре мозга при подготовке и выполнении движения, и хронометрировать эти процессы, т.е. установить временные границы их протекания.
Одним из важных направлений в исследовании психофизиологии двигательного акта является изучение комплекса колебаний потенциалов мозга, связанных с движениями (ПМСД). Значение этого феномена для понимания физиологических механизмов организации движения очень велико, потому что изучение ПМСД позволяет выявить скрытую последовательность процессов, происходящих в коре мозга при подготовке и выполнении движения, и хронометрировать эти процессы, т.е. установить временные границы их протекания.
Компонентный состав ПМСД. Впервые этот комплекс, отражающий процессы подготовки, выполнения и оценки движения был зарегистрирован в 60-е годы. Оказалось, что движению предшествует медленное отрицательное колебание — потенциал готовности (ПГ). Он начинает развиваться за 1,5 - 0,5 с до начала движения. Этот компонент регистрируется преимущественно в центральных и лобно-центральных отведениях обоих полушарий. За 500-300 мс до начала движения ПГ становится асимметричным — его максимальная амплитуда наблюдается в прецентральной области, контралатеральной движению. Примерно у половины взрослых испытуемых на фоне этого медленного отрицательного колебания незадолго до начала движения регистрируется небольшой по амплитуде положительный компонент. Он получил название "премоторная позитивность" (ПМП). Следующее по порядку быстро нарастающее по амплитуде отрицательное колебание, так называемый моторный потенциал (МП), начинает развиваться за 150 мс до начала движения и достигает максимальной амплитуды над областью моторного представительства движущейся конечности в коре головного мозга. Завершается этот комплекс потенциалов положительным компонентом примерно через 200 мс после начала движения.
Функциональное значение компонентов. Принято считать, что потенциал готовности (ПГ) возникает в моторной коре и связан с процессами планирования и подготовки движения. Он относится к классу медленных отрицательных колебаний потенциала мозга, возникновение которых объясняют активацией нейрональных элементов соответствующих участков коры.
Гипотезы относительно функционального значения ПМП различны.
Это колебание рассматривают и как отражение подачи центральной команды от коры к мышцам, и как результат релаксации коры после завершения определенного этапа организации движения, и как отражение процессов подавления ассоциированных движений другой конечности, и как обратную связь от мышечных афферентов. В настоящее время некоторые авторы полагают, что ПМП являются лишь отражением начала моторного потенциала.
При регистрации МП у обезьян в составе МП были выделены два субкомпонента. Первый субкомпонент соотносят с активацией моторной коры, связанной с инициацией движения (синаптическая активность пирамидных нейронов), а второй — с активацией полей 2.3 и 4 по Бродману. Регистрация МП у человека, больного эпилепсией, позволила выделить в нем три компонента. Первый компонент был назван потенциалом инициации. Он имеет высокую амплитуду и возникает после начала движения в прецентральной контралатеральной коре. Второй, возникающий после начала миограммы, более локализованный в контралатеральном соматосенсорном поле может быть связан как с инициацией движения, так и с сенсорной обратной связью. Третий компонент отражает импульсацию, поступающую с мышечных афферентов в кору.
Следующий за МП позитивный потенциал рассматривается как отражение обратной афферентации, поступающей с периферических рецепторов, восходящей активности от моторных центров, операции сравнения между моторной программой и нейрональной картиной ее исполнения или процессов релаксации коры после выполнения движения.
Волна ожидания. Помимо ПМСД, описан еще один электрофизиологический феномен, который по своей сути близок потенциалу готовности. Речь идет об отрицательном колебании потенциала, регистрируемого в передних отделах коры мозга в период между действием предупреждающего и пускового (требующего реакции) сигналов. Это колебание имеет ряд названий: волна ожидания, Е-волна, условное негативное отклонение (УНВ). Е-волна возникает через 500 мс после предупредительного сигнала, ее длительность растет с увеличением интервала между первым и вторым стимулами. Амплитуда Е-волны растет прямо пропорционально скорости двигательной реакции на пусковой стимул. Она увеличивается при напряжении внимания и увеличении волевого усилия, что свидетельствует о связи этого электрофизиологического явления с механизмами произвольной регуляции двигательной активности и поведения в целом.
Нейронна активність та нейронні механізми поведінки.
Нейронная активность
Функциональные кортикальные колонки. В моторной зоне коры у человека имеются так называемые гигантские пирамидные клетки Беца, которые организованы в обособленные колонки. Пирамидные клетки, выполняющие сходные функции, расположены рядом друг с другом, иначе было бы трудно объяснить точную соматотопическую организацию коры. Такие двигательные колонки способны возбуждать или тормозить группу функционально однородных мотонейронов.
Регистрация активности одиночных пирамидных клеток с помощью вживленных микроэлектродов у животных, выполняющих различные движения, позволила установить принципиально важный факт. Нейроны коры, регулирующие деятельность какой-либо мышцы, не сосредоточены в пределах только одной колонки. Двигательная колонка в значительной степени представляет собой функциональное объединение нейронов, регулирующих деятельность нескольких мышц, действующих на тот или иной сустав. Таким образом, в колонках пирамидных нейронов моторной коры представлены не столько мышцы, сколько движения.
Нейронные коды моторных программ. Кодирование информации в нейроне осуществляется частотой его разрядов. Анализ импульсной активности нейронов при выработке у животных различных моторных программ показал, что в их построении участвуют нейроны разных отделов двигательной системы, выполняя при этом специфические функции. По некоторым представлениям включение моторных программ происходит благодаря активации так называемых командных нейронов. Командные нейроны находятся, в свою очередь, под контролем высших корковых центров. Торможение командного нейрона приводит к остановке контролируемой им программы, возбуждение, напротив, к активизации нервной цепи и актуализации моторной программы.
Вовлечение командных нейронов в целостную деятельность мозга определяется текущей мотивацией и конкретной двигательной программой направленной на удовлетворение этой мотивации. Двигательная программа, чтобы носить приспособительный характер, должна учитывать все сигнально значимые компоненты внешней среды, относительно которых совершается целенаправленное движение, т.е. строиться на принципе мультисенсорной конвергенции.
Пропріоцепція.
ПРОПРИОЦЕПЦИЯ
Для успешной реализации движений необходимо, чтобы управляющие этими движениями центры в любой момент времени располагали информацией о положении звеньев тела в пространстве и о том, как протекает движение. В то же время движения являются мощным средством получения информации об окружающем мире. Некоторые виды сенсорной информации, например осязательная (гаптическое чувство) и зрительная, вообще могут быть получены только посредством определенных движений (соответственно, кисти и пальцев или глаз). Таким образом, связь между сенсорикой и моторикой очень тесна. По образному выражению Н.А. Бернштейна, «в организме все моторы осенсорены, а сенсоры омоторены».
Особое значение для управления движениями имеют сигналы двух типов мышечных рецепторов – мышечных веретен и сухожильных органов Гольджи. В каждой мышце человека можно встретить группы более тонких и коротких, чем остальные, мышечных волокон, заключенных в соединительнотканную капсулу длиной в несколько миллиметров и толщиной в несколько десятков микрон. Из-за своей формы эти образования получили название «мышечные веретена», а заключенные в капсулу мышечные волокна называются «интрафузальными» (внутриверетенными).
Мышечные веретена – это сложные образования, имеющие как афферентную, так и эфферентную иннервацию. Толстое афферентное волокно группы Iа, проникая внутрь капсулы веретена, ветвится, и его окончания обвивают в виде спиралей центральную часть интрафузальных волокон. Эти окончания называют первичными. Многие веретена иннервируются также одним или несколькими волокнами группы II, а их окончания располагаются к периферии от первичных окончаний и называются вторичными окончаниями.
Оба типа окончаний механочувствительны и активируются при растяжении мышцы. При этом частота импульсов, поступающих в мозг от первичных окончаний, зависит от амплитуды и скорости растяжения, а вторичные окончания чувстительны лишь к величине растяжения. Чувствительность афферентов Iа и II может регулироваться путем изменения жесткости интрафузальных мышечных волокон. Такие изменения происходят под влиянием тонких (группа g) эфферентных двигательных волокон, идущих к веретену и являющихся аксонами g-мотонейронов. Различают два вида g- волокон, которые могут изменять чувствительность афферентов к величине растяжения и к скорости независимо (соответственно g-статические и g-динамические волокна).
В отличие от веретен, расположенных параллельно мышечным волокнам, сухожильные органы Гольджи располагаются последовательно в месте перехода мышечных волокон в сухожилие. Эти рецепторы являются специализированными окончаниями толстых афферентных волокон первой группы (Ib), и частота их разрядов пропорциональна развиваемой мышцей силе.
В суставных капсулах, внутрисуставных и внесуставных связках имеются механорецепторы типа Руффини, активирующиеся при движениях в суставе, главным образом вблизи его крайних положений. В мышце также очень много свободных нервных окончаний (группы III и IV). Все перечисленные ранее типы рецепторов обеспечивают так называемую «проприоцептивную чувствительность», снабжая ЦНС информацией о состоянии опорно-двигательного аппарата. Информацию о состоянии собственного тела могут давать также и другие виды рецепторов, формально не относящихся к проприоцептивным (рецепторы глубокой чувствительности, кожные рецепторы в области суставов и т.д.).
Управління рухами.
|
© 2013 wikipage.com.ua - Дякуємо за посилання на wikipage.com.ua | Контакти |