Глава 2. Восприятие электромагнитных волн: зрение
|
|
Адекватным стимулом для сетчатки глаза служит видимый спектр электромагнитных волн в пределах от 400 до 800 нм. В этом диапазоне различные длины волн воспринимаются как разные цвета.
В ответ на световые стимулы рецепторные клетки сетчатки посылают серии периодических импульсов в центральную нервную систему. Проследим этот процесс от сетчатки до зрительной коры.
Воспринимающий конец пути - сетчатка
Строение сетчатки. Сетчатка построена очень сложно. Она состоит из десяти слоев, и ее можно считать аванпостом нервных центров, так как большая часть ее ткани состоит из нервных клеток. В этом отношении она отличается от всех остальных органов чувств. Для восприятия имеют значение три типа клеток сетчатки.
Первый тип - это рецепторные клетки, палочки и колбочки. В центральной ямке, которая лежит в центре желтого пятна напротив зрачка, палочек нет, но к периферии их становится больше, чем колбочек. Сетчатка человеческого глаза содержит около 120 млн палочек и 6 млн колбочек. Примечательно, что свет, прежде чем достичь светочувствительного аппарата, должен пройти не только через оптическую систему глаза (роговицу, переднюю камеру глаза, зрачок, хрусталик, стекловидное тело), но также через слои сетчатки. Палочки и колбочки не связаны прямо со зрительным нервом, идущим в головной мозг иниооразуют синапсы с биполярными клетками, составляющими целый нейронный слой. Одна рецепторная клетка может быть соединена с несколькими биполярными, и, наоборот, несколько рецепторных клеток могут образовать синапсы с одной биполярной клеткой Это делает возможным широкий диапазон вариаций. В свою очередь биполяры подходят к ганглиозным клеткам, волокна которых образуют зрительный нерв. Связи между биполярными и ганглиозными клетками тоже многообразны. Совокупность палочек и/или колбочек, связанных со зрительным нервом через одну ганглиозную клетку (через посредство биполяров), называется рецептивным полем этой клетки Импульсы проводятся в мозг по волокнам примерно миллиона ганглиозных клеток приблизительно от 130 млн рецепторных клеток Поскольку отношение здесь составляет около 130 : 1, имеет место сильно выраженная конвергенция.
Острота зрения. Давно известно, что при нормальной оптической системе глаза у человека острота зрения составляет около 1 минуты; это значит, что глаз видит две точки раздельно, если расстояние между ними в центральной ямке, т е. в точке самого ясного видения, не меньше 1 дуговой минуты. Показано, что в этом случае между двумя стимулируемыми колбочками должна лежать одна "молчащая" колбочка, что соответствует расстоянию на сетчатке в 4 мкм (средний диаметр колбочек равен 3 мкм).
On- и off-клетки. Недавно установлено, что отдельные англиозные клетки при стимуляции сетчатки ведут себя по-разному. Куффлер (Kuffler) измерял потенциал действия ганглиозных клеток, вводя микроэлектроды в глаза разных млекопитающих животных. Он нашел, что в одних клетках световой стимул усиливал потенциал покоя, а в других снижал его (наблюдалась ритмическая спонтанная импульсация нестимулируемых рецепторных клеток). Это значит, что световой стимул включал активность одних клеток и выключал ее в других клетках… Соответственно эти два типа клеток называются on- и off-элементамн. Примечательно, что оп-элементы всегда окружены кольцом из off-элементов, и наоборот… Такое мозаичное распределение возбуждающих и тормозящих элементов в сетчатке обеспечивает первичную переработку информации в глазу. При засвете двух on-элементов их импульсы суммируются, прежде чем пойдет по зрительному пути. Но если одно световое пятнышко подействует на on-клетку, а другое - на off-клетку, два импульса, вызывающие противоположные эффекты, нейтрализуют друг друга, и может случиться, что мозг не получит никакой информации.
Чувствительность к контрастам. Импульсация при диффузном освещении всей сетчатки гораздо слабее, чем при освещении небольших ее участков Вероятно, это объясняется резко раз-1раниченным мозаичным распределением on- и off-клеток в сетчатке и показывает, что зрительные нейроны более всего чувствительны к контрасту. Разрешающая способность сетчатки выше всего в центральной ямке, потому что здесь каждая из тесно расположенных колбочек может составлять отдельное рецептивное поле (т.е. соединяться с одной ганглиозной клеткой). Таким образом, острота зрения в центральной ямке составляет 1 минуту (4 мкм) в отличие от периферии, где диаметр одного рецептивного поля может быть равен нескольким миллиметрам, что соответствует остроте зрения в 3 градуса.
У некоторых млекопитающих (например, у кролика) частота передаваемых импульсов меняется в зависимости от направления движения света. Ни для человека, ни для высших млекопитающих животных, например для кошки или обезьяны, такая зависимость не доказана. Эти детекторы, чувствительные к движению, обнаружены только в головном мозгу.
Критическая частота слияния мельканий (к. ч. с.) Это явление связано с очень важным свойством сетчатки. Мелькающий свет воспринимается как непрерывный: выше определенной частоты мелькания сливаются. В ранних исследованиях к. ч. с.- частоты, при которой мелькания исчезают,- применялось вертящееся колесо, состоявшее из прозрачных и непрозрачных секторов, позади которого находился источник света. Теперь для этой цели служит стробоскоп с электронным управлением, создающий мелькания нужной частоты. Хотя к. ч. с - это корковая функция, в эффекте слияния несомненно участвует и сетчатка Установлено, что частота электрических ответов в ганглиозных клетках глаза у подопытных животных соответствует частоте световой стимуляции. Выше критической частоты ответ перестает быть аналоговым, и возникает только один ганглиозный потенциал. Отдельные рецепторные клетки имеют каждая свою собственную к. ч с. Высказано предположение, что слияние в мозгу основано на индивидуальных к ч. с рецепторных клеток сетчатки.
Явления, о которых до сих пор шла речь, никак не связаны с длиной волны света, т.е. с цветом. Чувствительность разных рецепторов в сетчатке к разным длинам волн различна, и поэтому идущие в сетчатке процессы имеют существенное значение для цветового зрения.
Теория двойственности. Физиологи, изучающие органы чувств, и офтальмологи давно поняли, что два типа рецепторных клеток в сетчатке, т.е. палочки и колбочки, различны по своим функциональным свойствам. Колбочки служат для острого видения при дневном свете и реагируют на разные длины волн.
Палочки же выполняют функцию отличения света от темноты и тем самым служат для ночного зрения и видения в сумеречном свете. Порог возбуждения палочек ниже, а благодаря особому фотохимическому процессу световая чувствительность сетчатки в темноте повышается. Это называют темповой адаптацией. В темноадаптированном глазу чувствительность сетчатки приближается к максимально возможной: самым низким порогом для палочек является один квант лучистой энергии света. Обладая такой фантастической световой чувствительностью, палочки в то же время не реагируют на различия в длине волны; различение цвета составляет функцию колбочек, у которых зато световая чувствительность гораздо ниже (минимальный порог равен 5-7 квантам). В центральной ямке имеются только колбочки, и здесь, как уже говорилось, рецептивное поле может состоять из одной колбочки. К периферии возрастает число палочек. Таким образом, зрение основано на совместной функции рецепторов двух типов на воспринимающем конце.
Фотохимические процессы. И в палочках, и в колбочках генерации рецепторного потенциала, ведущего к возникновению импульсации, предшествуют специфические фотохимические процессы. В палочках содержится вещество родопсин, называемое также зрительным пурпуром, которое расщепляется под действием света (для расщепления одной молекулы требуется один квант). Продукты расщепления играют какую-то роль в генерации рецепторного потенциала. Это процесс обратимый и продукты распада - ретиналь и опсин - в темноте снова соединяются в зрительный пурпур. Подобные вещества можно обнаружить и в колбочках, но здесь механизм их распада недостаточно ясен. Согласно Уолду (Wald), в трех разных группах колбочек имеются три вида светочувствительных пигментов. Одни колбочки содержат пигмент, чувствительный к зеленому свету, а другие чувствительны соответственно к красному и синему свету. Это подтверждает трехцветную (трихроматическую) теорию Юнга (Joung), предложенную им еще в 1801 году. Согласно этой теории, восприятие около 160 оттенков, которые способен различать человеческий глаз, является результатом одиночных или суммируемых ответов трех видов рецепторов из которых каждый чувствителен к определенной длине волны, т.е. к определенному цвету.
Доминаторы и модуляторы. Трехцветная теория была подтверждена опытами Гранита, который с помощью микроэлектродов регистрировал потенциалы действия ганглинозных клеток в сетчатке кошки. Он нашел, что и в темно-, и в светоадаптированной сетчатке большая часть клеток (главным образом палочки) чувствительна к широкому спектру длин волн и реагирует на такие стимулы импульсацией. На кривой чувствительности сетчатки доминирует этот единый ответ на все цвета, который Гранит назвал доминаторным ответом. Но имеется также небольшая группа ганглиозных клеток, отвечающих импульсацией только на один из трех основных цветов. Эти ганглиозные клетки, связанные с рецептивными полями из колбочек, были названы модуляторами. Они видоизменяют импульсацию доминаторов в соответствии с длиной волны воспринимаемого света.
Как мы видели, сетчатка является важным звеном свето-воспринимающего аппарата. Здесь отбираются и группируются импульсы, возникающие в ответ на световые стимулы видимого спектра, топографически организуются контуры и модулируется восприятие цвета до того, как оно будет передано в мозг. 130 млн. рецепторных элементов связаны с одним миллионом ганглиозных клеток и соответствующих аксонов. Все это элементы сложной кодирующей системы, доставляющей информацию в еще более сложную передающую и декодирующую систему.
Таламус - центральная переключательная станция.
Выйдя из сетчатки, волокна ганглиозных клеток соединяются в зрительный нерв - зрительный путь, идущий к таламусу. Около ствола мозга половина волокон зрительного пути перекрещивается. Таким образом, оба больших полушария получают волокна от обеих сетчаток. Таламус, представляющий собой крупную массу нейронов, служит переключательной станцией для всех сенсорных путей, за исключением обонятельного нерва. Волокна зрительного пути образуют синапсы с клетками латерального коленчатого тела, лежащего в заднем отделе таламуса. С помощью метода вызванных потенциалов (см. выше) были составлены карты проекций сетчатки на латеральное коленчатое тело и установлено, что каждый участок сетчатки представлен соответствующей точкой в таламусе.
Какую роль в зрительном восприятии играют таламические нейроны, неизвестно. Животные с удаленной корой больших полушарий, сохранившие только таламус, не утрачивают элементарной способности к восприятию света, но не в состоянии распознавать и дифференцировать стимулы (предметы, пищу).
On- и off-поля. При микроэлектродном исследовании таламических клеток обнаружены on- и off-клетки, расположенные так же, как в сетчатке, с той разницей, что on-элементы еще резче отграничены от окружающих off-элементов. Таким образом, на этом уровне зрительной системы возможность отличать диффузное освещение от четких контуров еще более очевидна.
Каково общее значение таламуса для восприятия, неизвестно. Некоторые исследователи считают, что он играет важную роль в субъективной эмоциональной окраске сенсорных импульсов. Согласно этой теории, впервые предложенной Кэнноном (Cannon) в начале 20-го века, удовольствие или неудовольствие, получаемое от стимулов, зависит главным образом от таламуса. Тесная связь таламуса с сенсорной корой (см. ниже) указывает на единство таламокортикальной сенсорной функции.
Декодирующий центр в коре.
Среди отделов коры большого мозга, которые первыми стали известны физиологам, были зрительные области. Дженнари (Jennari) в 1776 г. описал зрительную кору. Теперь ее размеры и строение на макро- и микроуровне хорошо изучены. Выяснены также некоторые физиологические свойства зрительной коры, но декодирующий процесс как целое, т.е. сложная функция зрительного восприятия, полностью еще не раскрыт. Однако те немногие детали этого сложного механизма, которые стали известны в последние годы (главным образом благодаря обширным микрофизиологическим исследованиям), обнадеживают и побуждают к дальнейшему изучению.
Зрительная кора. При изучении корковой проекционной зоны сетчатки применялись почти все методы исследования, упомянутые в гл. 1. У обезьян эта зона обнаружена в заднем отделе затылочной доли; анатомические, нейрохирургические и неврологические данные говорят о том, что в головном мозгу человека она занимает то же положение. Центральная ямка представлена участком в несколько квадратных сантиметров на наружной, выпуклой поверхности затылочной доли. При стимуляции одной точки сетчатки светом или толчками электрического тока вызванные потенциалы возникают в нескольких, по меньшей мере в двух, точках коры.
В отличие от сравнительно однородной организации латерального коленчатого тела зрительная кора состоит из нескольких слоев. Волокна от коленчатого тела оканчиваются в четвертом (от поверхности коры) из шести различимых под микроскопом слоев. Обширная сеть волокон передает отсюда информацию к соседним слоям. От третьего п пятою слоев отходит множество волокон, идущих к подкорковым нейронам и соседним областям коры.
Примечательная особенность этой системы состоит в том, что вертикальных, колончатых связей между отдельными слоями гораздо больше, чем горизонтальных, боковых связей. Из такой организации следует, что корковыми проекциями отдельных рецептивных полей служат ограниченные вертикальные колонки. Эта колончатая организация была подтверждена электрофизиологическими данными.
Ориентационные колонки. Недавно Хьюбел и Визель (Hubel, Wiesel) установили, что все нейроны этих кортикальных колонок отвечают на простые линейные стимулы одного типа, т.е. на прямые световые линии (щели или границы контраста), идущие в определенном направлении. Например, если кошке показать вертикальную линию на белом поле, то от соответствующих клеток коры отводятся высокочастотные потенциалы действия.
Но если ориентацию линии изменить, электрический ответ этих клеток исчезнет. Клетки других колонок реагируют на линии с иной ориентацией. На этом основании в качестве основной аналитической единицы постулирован набор ориентационных колонок, содержащих все специальные разновидности в пределах 180° (гиперколонка). В то же время оказалось, что сложность ответов варьирует с расстоянием от поверхности коры, что указывает на существование нескольких слоев "простых" и более "сложных" клеток.
"Простые" и "сложные" клетки. Нейроны, отвечающие на простые линейные стимулы (щели, края или темные полосы), получили название "простых", а те, которые отвечают на стимулы сложной конфигурации и на движущиеся стимулы, были названы "сложными".
On- и off-клетки тоже можно обнаружить в зрительной коре, но здесь они расположены не концентрически. Эти два вида клеток резко разграничены, и направление разделяющей их границы зависит от ориентационной специфичности нейрона. Предполагается, что функция ориентационных колонок осуществляется путем латерального торможения.
Распределение колончатых функциональных единиц в зрительной коре неоднотипно. Показано, что "простые" клетки преобладают в поле 17 по Бродману, которое лежит в центре зрительной коры, а в полях 18 и 19 чаще встречаются "сложные" клетки. Высказано предположение, что информация, проанализированная и переработанная простыми клетками, передается сложным для дальнейшего анализа. Таким образом, весьма вероятно, что зрительное восприятие - результат кооперации разных клеток.
Информация, закодированная, проанализированная и переработанная в сетчатке и таламусе, декодируется несколькими миллионами нейронов в коре. В конечном результате, таким образом, получается аналоговое сообщение в соответствии с принципом отражения внешней среды.
"Сверхсложные" нейроны. В зрительной коре описан также третий класс нейронов, а именно "сверхсложные" клетки. Это истинные интегрирующие единицы, выполняющие в зрительной системе функцию синтеза. Они получают информацию от нижележащих нейронов и обеспечивают единообразие восприятия пространства и формы.
Восприятие цвета. Хотя эти электрофизиологические данные создают прочную основу для дальнейшего изучения работы мозга, их недостаточно для интерпретации восприятия в понятиях психологии. Множество вопросов все еще остается без ответа. Как на сравнительно простой пример укажем на то, что мы почти ничего не знаем о цветовоспринимающей функции мозга. Кажется несомненным, что сообщение о цвете кодируется доминаторно-модуляторной системой сетчатки, но нет никаких данных о перерабатывающей и декодирующей активности в латеральном коленчатом теле и коре. В таламусе обезьяны были исследованы нейроны разных типов, отвечающие на различные цветовые стимулы, но полученные результаты противоречивы. Скудность данных объясняется тем, что лабораторные млекопитающие (грызуны, кошки, обезьяны) плохо различают цвета. Собака, например, совсем их не различает. Большая часть информации получена в опытах на лягушках. И хотя эти опыты выполнены весьма остроумно и имеют большое значение, их результаты нельзя перенести на человека.
Центральная регуляция зрения.
Взаимодействие афферентных и эфферентных механизмов в зрительной системе очевиднее, чем в любой другой сенсорной системе. Для возникновения зрительного ощущения необходимы как сенсорные волокна, идущие от сетчатки в мозг, так и моторные, проводящие импульсы из мозга в глаз. Зрение невозможно без центрального контроля, т.е. без того, чтобы движения периферического сенсорного аппарата регулировались мозгом. Известны три вида эфферентных механизмов.
Зрачковый рефлекс. Самым известным примером регулирующей функции мозга служит зрачковый рефлекс на свет. Сенсорные импульсы проводятся афферентным звеном рефлекторной дуги - особыми волокнами зрительного нерва, идущего от сетчатки в мозг. Волокна этого типа образуют отдельные пучки в стволе мозга и идут к специальной группе мезэнцефальных нейронов (к ядру Эдингера - Вестфаля), где вступают в связь с эфферентными нервами, регулирующими величину зрачков в соответствии с интенсивностью и отдаленностью источника света Таким образом, стимул сам определяет размеры зрачка и количество света, которое сможет попасть в глаз в следующую тысячную секунды. Зрачковый рефлекс служит важным средством устранения "шума" из зрительной информации, приходящей извне. Блокада рефлекса атропином или каким-либо другим веществом, парализующим мышцы, которые сужают зрачок, снижает остроту зрения, а также нарушает восприятие формы и цвета.
Движения глаз. Работа мышц, движущих глазные яблоки, тоже подчинена центральной регуляции. Без этого зрительное восприятие было бы невозможно. Когда мы смотрим на предмет, наши глаза совершают безостановочные движения. При неподвижной сетчатке изображение не воспринимается; для того чтобы возникло зрительное восприятие, сетчатка должна смещаться относительно рассматриваемого объекта. Движения глазного яблока фотографическим путем регистрировал Ярбус, Притчард (Pritchard) доказал, что зрительное восприятие невозможно, если глазное яблоко не может перемещаться по отношению к стимулу. Он укрепил микропроектор на контактной линзе и таким способом устранил движение сетчатки относительно изображений, которые проецировались на нее с помощью этого аппарата. В первую секунду опыта испытуемый видел ясное изображение, но потом оно расплывалось, исчезало и восстанавливалось лишь частично.
Три пары мышц, приводящих в движение глазное яблоко, находятся фактически под контролем затылочной зрительной коры, но, кроме того, ими непосредственно управляет поле 8 по Бродману в премоторной зоне лобной коры (или, точнее, ядра среднего и продолговатого мозга, подчиненные полю 8.
Эфферентный контроль сетчатки. О третьем важном механизме центральной регуляции зрительного восприятия мало что известно. Как уже говорилось, установлено, что ко всем органам чувств от головного мозга идут эфферентные волокна, которые, по всей вероятности, проводят импульсы центрального контроля. Этот эфферентный контроль сетчатки описан Додтом (Dodt), а также Гранитом. Между биполярными и ганглиозными клетками сетчатки лежит слой так называемых амакриновых клеток. Аксоны этих клеток образуют синапсы с волокнами, которые оканчиваются в неидентифицированных областях центральной нервной системы, скорее всего в ретикулярной формации (см. ниже). Вызываемые световым стимулом импульсы в рецепторах можно модифицировать, раздражая эти волокна.
Восприятие формы.
Если перед самым глазом испытуемого поместить освещенную внутреннюю поверхность полусферы, например половинку шарика для пинг-понга, то будет виден лишь слабый свет без какого-либо специфического зрительного восприятия. Это отсутствие перцептивного акта не зависит от цвета и освещенности однородного поля и связано с отсутствием минимального расчленения. Для того чтобы возникло зрительное восприятие, объект должен обладать некоторой организацией и структурой (разумеется, нужны еще упомянутые выше движения глаза, которые тоже служат необходимым условием зрительной функции). Отсутствие расчленения раньше или позже приводит к галлюцинациям, например к миражам, которые люди иногда видят на плоских равнинах, где однотонность ландшафта и неба вызывает галлюцинации.
Восприятие формы. Восприятие форм и структур является врожденной способностью, поскольку описанные выше механизмы обработки информации в сетчатке, таламусе и коре действуют с момента рождения. Тем не менее в восприятии формы играют также очень важную роль элементы, приобретаемые в процессе научения. Показано, что дети в возрасте до шести лет не распознают загадочные картинки или спрятанные фигуры, а также не разделяют фотографии, наложенные одна на другую. Это объясняется их ограниченной способностью к восприятию форм. Детям нужно больше времени, чтобы распознать сложные изображения, но в то же время перевернутые картинки они узнают с большей легкостью. До шестилетнего возраста зрение у детей туннельное, т.е. фигуры, лежащие на периферии поля зрения, не воспринимаются. Даже у младенцев внимание гораздо легче привлечь изображениями знакомых, чем незнакомых предметов… Все эти факты говорят о роли опыта и научения в восприятии форм. После удаления врожденных катаракт нужны месяцы, чтобы зрительные впечатления стали соответствовать прежнему тактильному опыту, и функция восприятия форм развивается лишь постепенно. Цвета такие больные начинают различать раньше, чем формы. Вначале они могут не отличить квадрат от шестиугольника или петушиные перья от конского хвоста. Таким образом, эти наблюдения опять-таки свидетельствуют о роли факторов научения в восприятии форм.
Восприятие форм у животных. Функция восприятия паттернов обнаружена и у некоторых беспозвоночных животных. Так, например, осьминог различает разные треугольники, но не способен дифференцировать зеркальные изображения. Интересно отметить, что он лучше отличает горизонтальные линии от вертикальных или фигуру  от
от  , чем
, чем  от
от 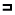 . Некоторые виды беспозвоночных (например, прыгающие пауки) узнают свои замаскированные безжизненные жертвы. Насекомые со сложными глазами обычно хорошо различают формы. Поразительную способность пчел к восприятию цвета и форм уже давно описал Фриш (Frisch). Этой способностью обладают и почти все виды позвоночных животных. Всем известно зрительное чувство направления у перелетных птиц. Например, славка находит свой путь ночью по положению звезд, и ее можно также стимулировать, проецируя перед ней в планетарии световые пятнышки, изображающие звезды. Восприятие форм описано Лэшли (Lashley) у крыс и Клювером (Kluver) у обезьян.
. Некоторые виды беспозвоночных (например, прыгающие пауки) узнают свои замаскированные безжизненные жертвы. Насекомые со сложными глазами обычно хорошо различают формы. Поразительную способность пчел к восприятию цвета и форм уже давно описал Фриш (Frisch). Этой способностью обладают и почти все виды позвоночных животных. Всем известно зрительное чувство направления у перелетных птиц. Например, славка находит свой путь ночью по положению звезд, и ее можно также стимулировать, проецируя перед ней в планетарии световые пятнышки, изображающие звезды. Восприятие форм описано Лэшли (Lashley) у крыс и Клювером (Kluver) у обезьян.
Зрительное восприятие глубины и расстояния.
Отдаленность предмета человек может оценивать, даже если он смотрит одним глазом. При монокулярном зрении расстояние различных предметов от глаза, т.е. третье измерение, выводится из разных признаков - относительной величины предметов, их освещенности и взаимного положения. Эта особенность приобретается в раннем детстве условнорефлекторно.
В одном опыте клетку с только что вылупившимися цыплятами, которые обладают монокулярным зрением, освещали снизу, после того как на дно клетки было насыпано зерно. Через несколько дней цыплятам стали показывать фотографии клетки, освещенной снизу, и они принимались клевать эти картинки Это показывает, что научение делает возможной оценку расстояния и глубины даже при монокулярном зрении.
Стереоскопическое зрение. Восприятие глубины более совершенно при бинокулярном зрении. При этом изображения, информация о которых передается от обоих глаз в кору, не совсем одинаковы. Чем ближе предмет к глазам, тем больше различие (диспаратность) между изображениями. На слиянии обоих изображений основано объемное, т.е. стереоскопическое, зрение.
В изображении на правой сетчатке больше видна правая сторона предмета, а на левой сетчатке - левая сторона, как если бы два фотографа снимали двумя камерами, расположенными на небольшом расстоянии одна от другой. В зрительном центре в коре оба изображения сливаются, создавая глубинный эффект бинокулярного зрения. Он возникает вследствие стимуляции неидентичных точек сетчатки. Элементы объектов, которые проецируются на неидептичные точки и сдвинуты к височным сторонам сетчаток, воспринимаются как более близкие, а элементы, сдвинутые на сетчатках ближе к носу, - как более удаленные. Элементы, попадающие на идентичные точки обеих сетчаток, видны в одной и той же плоскости.
Две плоские картинки можно зрительно совместить при помощи зеркал или призм, расположенных таким образом, что каждый глаз будет видеть лишь одну из этих картинок, и в результате возникнет впечатление объемного восприятия. Применяемый с этой целью прибор называется стереоскопом. При рассмотрении таким способом двух идентичных картинок эффект глубины не возникает. Стереоскоп приспособили также для рассмотрения отдаленных предметов, объемное восприятие которых невозможно из-за большого расстояния. В созданном таким путем телестереоскопе диспаратность между двумя глазами усиливается благодаря системе зеркал.
Подделанные копии документов или фальшивые деньги, помещенные вместе с оригиналом в стереоскоп, создают объемное изображение, и тем самым обнаруживается подделка, так как выявляются небольшие различия между двумя изображениями.
Слияние. Стереоскопическое зрение представляет собой кортикальную функцию, приобретаемую в результате научения. Бинокулярные изображения сливаются благодаря иптегративной деятельности клеток коры. Микроэлектродными исследованиями в коре мозга обнаружены нейроны, отвечающие на стимуляцию обеих сетчаток, что говорит о конвергенции на этих нейронах импульсов, приходящих от обоих глаз.
По зрительному восприятию накоплено больше экспериментальных данных, чем по всем остальным органам чувств, взятым вместе. У человека зрительная система - наиболее высокоорганизованная из сенсорных систем, чтобы это понять, не нужно быть биологом или психологом очевидно, что зрение доминирует над всеми органами чувств. Поэтому остальные сенсорные системы мы рассмотрим только в плане их отличия от зрительной.
Глава 3.
Восприятие механических колебаний: слух
Согласно шеррингтоновской классификации, слуховой аппарат принадлежит к классу механорецепторов Это система специализированная для приема механических колебаний частотой от 16 до 20000 Гц (полных колебаний в секунду) Переработка информации в слуховом аппарате и в зрительной системе во многом сходна.
Строение слухового анализатора
Общие особенности. Слуховой аппарат человека - весьма чувствительный рецептор Его строение тщательно исследуют представители новой дисциплины - бионики, которые хотели бы создать прибор, обладающий такой же точностью и избирательностью Чувствительность человеческого уха всем хорошо известна. Ухо реагирует на чрезвычайно слабые колебания почти эквивалентные по энергии ударам молекул воздуха о барабанную перепонку. В то же время оно противостоит чрезвычайно сильным вибрациям, создаваемым пневматическими сверлами или сверхзвуковыми самолетами. В качестве примера избирательности можно указать на всем знакомую ситуацию, когда в шумном помещении человек способен сосредоточить внимание на голосе одного говорящего Точно так же музыканты отбирают из звучания целого оркестра мелодию исполняемую на одном инструменте, и следуют за ней.
Периферический рецепторный орган слуха - кортиев орган - находится во внутреннем ухе. Он принимает колебания которые передаются к нему барабанной перепонкой и косточками среднего уха, преобразует их в электрические импульсы и направляет в мозг. Ухо человека нечувствительно к колебаниям частотой менее 16 Гц, и поэтому звуки, производимые движениями его собственных мышц, сухожилий и других частей тела обычно не слышны. Самые высокие частоты, граничащие с ультразвуком, воспринимают только молодые люди; верхний предел слышимых частот после 30 лет постепенно снижается на 100-120 Гц в год.
Роль среднего уха. Звуковые волны колеблют барабанную перепонку, и эти колебания передаются через среднее ухо по трем маленьким косточкам (молоточку, наковальне и стремечку) жидкости, наполняющей внутреннее ухо. Самая последняя из трех косточек - стремечко (весом около 1,2 мг) соприкасается с этой жидкостью через овальное окно и, действуя подобно поршню, приводит ее в движение в соответствии с ритмом звуковых волн. В свою очередь колебания жидкости заставляют колебаться мембрану кортиева органа. Эта сложная передающая система очень эффективна колебания большой амплитуды и малой энергии трансформируются в колебания гораздо меньшей амплитуды, но в 20 раз большей интенсивности.
Звуковые волны передаются не только через барабанную перепонку; кости черепа тоже способны проводить колебания. Звук нашего собственного голоса, каким мы его слышим сами, всегда отличен от того, каким его слышат другие, так как звуковые волны, образующиеся в гортани и ротовой полости, доходят до внутреннего уха также путем костной проводимости. Таким образом, слышимый нами звук - результат двойной передачи и через среднее ухо, и через кость. А другие люди воспринимают те же звуки только через среднее ухо, менее чувствительное к низким частотам.
Кортиев орган. Этот орган содержит волосковые клетки; когда до них доходят колебания давления, передающиеся во внутреннее ухо, они генерируют рецепторные потенциалы. Колебания жидкости во внутреннем ухе передаются базилярной (основной) мембране, лежащей в улитке, которая имеет форму спирали из 23/4 витков общей длиной 33 мм. Низкочастотные колебания действуют на апикальные (верхушечные) участки базилярной мембраны, а высокие тоны - на ее основание. При этом волосковые клетки, деформируясь, подвергаются механической стимуляции и преобразуют аналоговый сигнал смещения в серию бинарных сигналов - импульсов.
Таламический центр. Каждая волосковая клетка соединена с отростками двух нейронов - биполярных нервных клеток, которые находятся в ганглиях, расположенных близ внутреннего уха. Это первые нейроны слухового пути, который в отличие от зрительного состоит не из трех, а из пяти нейронов. Большая часть этих волокон, но не все по пути в большой мозг перекрещиваются. Таламус является важной передаточной станцией: от лежащего в нем медиального коленчатого тела начинается пятый нейрон, который идет к слуховой коре.
Слуховая кора. Слуховая кора точно картирована с помощью метода вызванных потенциалов. У верхней границы височной доли, близ нижнего края сильвиевой борозды, обнаружено множественное представительство слуховой системы. Это значит, что слуховые импульсы, приходящие от отдельных клеток кортиева органа, активируют в коре одновременно несколько нейронных групп. Кортикальные проекционные области имеют такую же топическую структуру, как зрительная кора, т.е. импульсы, вызываемые высокими частотами, проецируются в одни точки, а вызываемые низкими частотами - в другие в соответствии с различиями в эффекте стимуляции разных участков кортиева органа. Эта так называемая тонотопическая организация характерна для всего слухового пути, т.е. для улитки, медиального коленчатого тела и слуховой коры.
Кодирование и декодирование в слуховой системе.
Серии потенциалов действия в биполярных нейронах, которые контактируют с клетками, реагирующими на звуковые стимулы, изучаются при помощи микроэлектродной методики. В некоторых клетках импульсы могут возникать и без стимуляции (нейроны с фоновой частотой 10 и 50 Гц). Клетки второго типа называют "молчащими", так как они генерируют импульсы только в ответ на звуковые стимулы.
Отдельные звуки вызывают импульсы в ограниченной группе волокон слухового пути; эти волокна окружены "молчащими", заторможенными волокнами. Явление контраста (т.е. состояние, когда возбужденные группы клеток окружены заторможенными), о котором шла речь в связи со зрительным восприятием, встречается и в слуховом пути. Восприятие высоты тона (частоты колебаний) зависит от положения возбужденных волокон в слуховом пути или, точнее, от положения их границ с соседними волокнами. Что касается интенсивности, то она зависит от числа возбужденных волокон. Оба параметра играют важную роль при центральном декодировании.
Тонотопическую организацию слуховой коры соответственно частотам надо представлять себе как статистическую, а не как проекцию из точки в точку. Соседние области могут значительно перекрываться, но в то же время они могут очень различаться по своей частотной чувствительности. Колончатая организация, найденная в зрительной коре, обнаружена и здесь. Все участки коры, расположенные вертикально под нейронами, специфически реагирующими на звуки определенной частоты, отвечают на специфический стимул одинаково.
Восприятие акустической конфигурации и направления к источнику звука
Установлено, что кошки с удаленной корой сохраняют способность различать разные тоны. Следовательно, различение частоты является функцией таламуса, а кора, по-видимому, выполняет более сложные функции.
Снова слияние. Импульсы, вызываемые стимуляцией обоих ушей, объединяются в медиальном коленчатом теле и в клетках коры, подобно тому как импульсы, приходящие от обоих глаз, объединяются в зрительной коре. Слуховое ощущение является результатом активности нейронов, отвечающих на специфические частоты, а также вышележащих клеток с координирующей функцией, подобной функции "сложных" зрительных клето
|
© 2013 wikipage.com.ua - Дякуємо за посилання на wikipage.com.ua | Контакти |